Espelhos do genoma: retrocópias como doppelgängers moleculares
Na mitologia do doppeltgänger, do alemão “duplo ambulante”, as pessoas poderiam ter uma cópia de si mesmas, um gêmeo do mal, o qual estaria relacionado a presságios de má sorte. Há exemplos de doppelgänger na arte, assim como há testemunhos daqueles que afirmam, categoricamente, terem passado pela experiência de encontrar a própria contraparte na vida real.
O escritor alemão Goethe (2014), em sua obra “Os Sofrimentos do Jovem Werther”, retrata um episódio no qual o herói da trama, o jovem Werther, presencia, ao passear pelo campo, uma figura idêntica a si mesmo e trajando as mesmas roupas. Essa manifestação fantasmagórica foi interpretada de forma atônita pela personagem como um aviso de que o seu fim estava próximo.
Um dos casos mais famosos de aparição de doppeltgänger remete ao ex-presidente dos EUA Abraham Lincoln. Em 1865, poucos dias que antecederam o seu assassinato, Lincoln teria visto a própria imagem refletida numa janela do segundo andar de um edifício em Washington DC. A preocupação se instaurou de fato após presenciar novamente o seu clone em outras janelas durante o percurso que fazia. Diz, então, a lenda que o presidente teria relatado o ocorrido a sua esposa e amigos próximos e manifestado o temor que algo terrível estava prestes a suceder.
Não obstante a tradicional narrativa maligna, em algumas histórias modernas e contos, encontram-se exemplos de doppelgängers que agem positivamente. No romance “Lugar Nenhum” de Neil Gaiman (2016), o herói Richard Mayhew se encontra em uma Londres subterrânea fantástica onde, após um incidente, ele se vê praticamente esquecido por todos. Durante sua jornada, Richard encontra “reflexos” de si mesmo em personagens do submundo que acabam funcionando como espelhos, ajudando-o a encontrar sua verdadeira força e propósito.
Esse conceito também aparece em histórias de séries como Doctor Who, onde versões alternativas dos protagonistas às vezes agem como guias Esses exemplos reinterpretam o “gêmeo malvado” como um guardião que aparece em momentos de perigo, ajudando o protagonista a evitar acidentes ou encontrar o caminho certo. Em vez de ser uma sombra ameaçadora, o doppelgänger seria como um guia secreto, cuidando da personagem principal de longe e garantindo que ela se torne uma pessoa melhor, sem jamais ser descoberto como um “outro eu”. Essa versão transforma o mito, associando-o à proteção e autodescoberta.
Tal qual indivíduos estariam sujeitos a trombar pelas veredas da vida com o seu doppeltgänger, os genes, especificamente aqueles responsáveis pela codificação de proteínas, também estariam à mercê da mesma sorte. Mas então o que seria um doppeltgänger no contexto genômico?
Genes codificadores e L1
Os genes codificadores são aqueles genes transcritos a partir do genoma (nosso DNA), situado dentro do núcleo da célula, para moléculas de RNA mensageiro que, por sua vez, são transportadas para o citoplasma e lá encontram os ribossomos onde são traduzidas em proteínas. Durante essa trajetória, o RNA mensageiro passa por alguns percalços, podendo ser degradado por outras moléculas em resposta a sinais específicos antes mesmo de adentrar o território ribossomal (ALBERTS et al., 2017). Além de poder ser degradado, sobretudo após ser traduzido, existe a possibilidade de esse RNA mensageiro ser abduzido no citoplasma pelas proteínas de um outro gene, mais precisamente um elemento móvel (transponível) do genoma. O gene responsável por tal crime se chama LINE1 (ou apenas L1 para os íntimos) (KAESSMANN; VINCKENBOSCH; LONG, 2009).
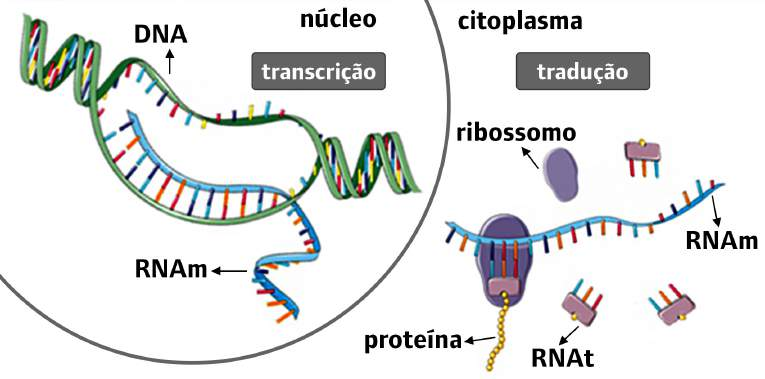
Fonte: (VASCONCELOS et al., 2021, fig. 4)
O LINE1, da abreviatura em inglês Long Interspersed Nuclear Element-1 (Elemento nuclear intercalado longo-1) é um gene que possui a capacidade de criar cópias de si mesmo e espalhá-las pelo genoma de seu hospedeiro. Ocorre que o L1 codifica a informação genética para a produção de pelo menos duas proteínas:
- a primeira localizada na Fase de Leitura Aberta 1, ORF1 (Open Reading Frame), quando traduzida dá origem a proteína ORF1p – a qual é uma proteína ligante de RNA (MARTIN, 2010);
- a segunda está localizada na Fase de Leitura Aberta 2, ORF2, e origina a proteína ORF2p – que por sua vez é uma proteína com atividades endonucleolítica (SULTANA et al., 2017) e transcriptase reversa (COST et al., 2002).
O habitual em humanos, e outros mamíferos, é o RNA mensageiro conter os códigos para a produção de uma única proteína (monocistrônico). Já aqui, temos um exemplo de RNA mensageiro policistrônico – ou mais precisamente, bicistrônico (por conta das duas proteínas ORF1p, ORF2p). Assim como os outros genes codificadores , o L1 é transcrito para o seu RNA mensageiro no núcleo e este, ao se transladar para o citoplasma, poderá ser traduzido nas proteínas supracitadas.
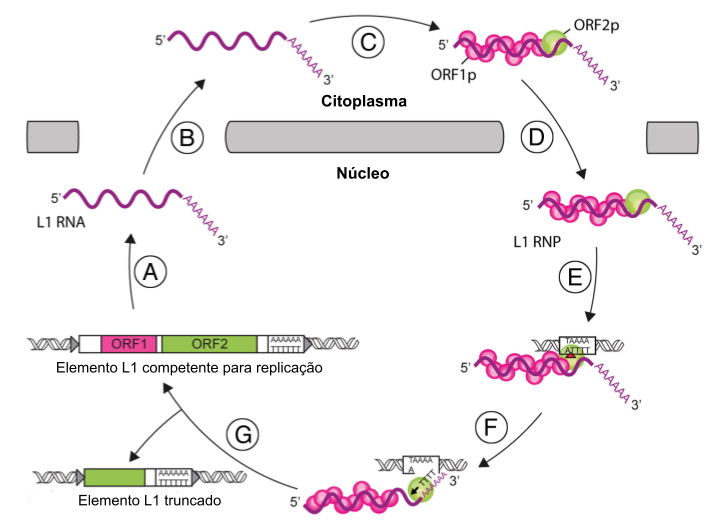
Fonte: (VIOLLET; MONOT; CRISTOFARI, 2014, fig. 1)
As proteínas ORF1p e ORF2p, após a sua síntese, conseguem interagir com o seu próprio RNA mensageiro no citoplasma, formando um complexo de moléculas de RNA e proteínas – chamado, portanto, de complexo ribonucleoproteico (WEI et al., 2001). Um complexo proteico funciona como uma matriz industrial, na qual cada setor é responsável por uma parte do processo de produção. Sendo assim, o setor da ORF1p é incubido da estabilização da molécula de RNA, protegendo-a de degradação e facilitando o acoplamento da ORF2p. O setor da ORF2p dá início às suas atividades após o deslocamento do nosso complexo ribonucleoproteico do citoplasma para o núcleo. No núcleo, a ORF2p identifica os padrões cromossomais necessários para a inserção de uma nova cópia de L1 e, ao encontrá-los, cliva a molécula de DNA, gerando uma abertura (atividade endonucleolítica). É nessa abertura que será inserido pela atividade de transcriptase reversa da ORF2p um clone do RNA mensageiro associado ao complexo (MARTIN, 2010; MARTIN et al., 2005). Essa outra atividade da ORF2p transcreve de RNA para DNA – até por isso é chamada de transcriptase reversa, haja vista que a transcriptase transcreve de DNA para RNA, então aqui a ORF2p realiza a reação reversa.
Os olhos mais atentos devem ter percebido a similaridade que há entre L1 e os retrovírus (como o HIV). Ambos utilizam uma transcriptase reversa para se integrar ao genoma do hospedeiro, mas fazem isso com objetivos diferentes em termos evolutivos. Os vírus têm ciclos de vida que envolvem a infecção de novas células e novos organismos, o que os torna parasitas completos (TELESNITSKY; GOFF, 1997), enquanto os LINE1 “viajam” apenas dentro de um organismo, inserindo-se em novos locais no DNA da célula hospedeira e nas células descendentes. Mesmo assim, ambos utilizam a maquinaria celular para sua replicação e dispersão. Esse comportamento “autônomo” alinha-se à ideia de Richard Dawkins (2007), no livro O Gene Egoísta, sobre a “sobrevivência do mais persistente”, em que elementos genéticos que garantem sua própria replicação são favorecidos, mesmo se isso tiver efeitos potencialmente danosos para o organismo hospedeiro.
Os clones de L1 podem ser íntegros, preservando as duas regiões de leitura aberta e, portanto, capazes de retrotranspor ativamente, gerando novas cópias de si mesmo. Contudo, muitas vezes os L1 inseridos são truncados, perdendo uma ou ambas as ORFs e, assim, tornando-se incapacidade de se mobilizar (SZAK et al., 2002). Curiosamente, algumas dessas cópias truncadas podem ser cooptadas pelo genoma do hospedeiro e acabar adquirindo novas funções regulatórias, atuando como elementos de controle gênico ou de expressão (HAN; BOEKE, 2005).
Outra questão a ser levada em consideração é o fato de as inserções de L1 poderem ocorrer em regiões genômicas altamente transcritas e, por conseguinte, coincidirem com a localização de outros genes, causando mutações prejudiciais, ou criando novidades genéticas potencialmente vantajosas. Há estudos que indicam que a atividade de L1 pode induzir mutações somáticas que resultam na origem de câncer: como alguns casos de câncer de cólon (OGINO et al., 2008), mama (VAN HOESEL et al., 2012) e pulmão (SUN et al., 2022). Mas ao mesmo tempo que esse elemento pode causar doenças, ele também possui um papel na diversificação gênica. No cérebro, por exemplo, há evidências de que L1 está envolvido na geração de diversidade entre neurônios (GARZA et al., 2023), o que contribuiria para a complexidade do desenvolvimento cerebral e da plasticidade neural, sugerindo que a atividade controlada de L1 pode ter efeitos benéficos em contextos específicos, como a cognição e o aprendizado.
Retrocópia – o duplo ambulante de um gene codificador
O gene de LINE1, como visto, possui a capacidade ímpar de criar cópias de si mesmo e espalhá-las pelo genoma. Esse ato de se auto copiar é chamado de reação em cis, porém acontece também de a maquinaria de duplicação oriunda de LINE1, ocasionalmente, interseptar o RNA mensageiro de um outro gene e, consequentemente, gerar uma cópia dele, sendo esse ato nominado de reação em trans (KAESSMANN; VINCKENBOSCH; LONG, 2009). As cópias gênicas produzidas pela atividade de LINE1 são carinhosamente conhecidas como retrocópias.
Assim como um doppelgänger pode surgir inesperadamente na vida de uma pessoa, carregando sua imagem, mas sem sua essência completa, as retrocópias são duplicatas de genes codificadores que surgem no genoma de maneira fortuita. No entanto, diferentemente de uma duplicação convencional do DNA, em que tanto exons quanto íntrons são copiados, as retrocópias nascem desprovidas de íntrons e das sequências regulatórias que determinam sua correta expressão. Elas são, por assim dizer, sombras genéticas de seus genes ancestrais, versões destituídas de alguns componentes essenciais para sua função original (VANIN, 1985).
Portanto, muitas retrocópias acabam sendo silenciadas, tornando-se elementos fósseis do genoma, incapazes de se expressar (de serem transcritas) e relegadas ao esquecimento evolutivo, tornando-se, de fato, pseudogenes. No entanto, algumas poucas escapam a esse destino e, ao adquirirem novos elementos regulatórios, podem não apenas assumir funções inovadoras, mas também preservar a atividade do gene parental (KAESSMANN; VINCKENBOSCH; LONG, 2009). Em certos casos, a retrocópia mantém a mesma função do gene original, diferenciando-se principalmente no seu padrão de expressão, o que permite a especialização em tecidos ou condições biológicas específicas (BURKI; KAESSMANN, 2004). Além disso, algumas retrocópias atuam como reguladores gênicos, interferindo na expressão de outros genes por meio da produção de RNAs não codificantes ou mesmo pela influência sobre elementos regulatórios próximos (POLISENO et al., 2010). Dessa forma, enquanto algumas retrocópias se tornam genes “novos” e funcionalmente ativos, outras exercem papéis sutis, mas relevantes, na regulação da expressão gênica, sobretudo dos seus genes parentais (dos quais elas se originaram).
Curiosamente, assim como o mito dos doppelgängers apresenta narrativas que oscilam entre o presságio sombrio e a influência positiva, as retrocópias também carregam essa dualidade. Algumas inserções podem levar a mutações prejudiciais, interrompendo genes essenciais e contribuindo para o desenvolvimento de doenças, como o câncer. Outras, porém, podem conferir vantagens adaptativas, criando novidades genéticas, influenciando a expressão de redes gênicas e contribuindo para a origem de novas características e complexidades em diversos organismos.
Em última análise, tanto nos mitos quanto no genoma, os doppelgängers – sejam eles humanos ou moleculares – são reflexos inesperados da existência original. Enquanto alguns podem parecer meras sombras sem propósito, outros se tornam protagonistas de novas histórias, redefinindo seu papel no curso da evolução das espécies.
Epílogo
A analogia entre doppelgängers e retrocópias gênicas ilustra de maneira instigante a dualidade dessas cópias genéticas no genoma humano. Assim como um doppelgänger pode ser apenas uma réplica inofensiva ou assumir um papel significativo na vida de seu original, as retrocópias podem permanecer como pseudogenes silenciosos ou adquirir funções inovadoras, influenciando a evolução e a regulação da expressão gênica.
A atividade de LINE1 é a força motriz por trás da geração dessas cópias gênicas, tornando o genoma um ambiente dinâmico e sujeito a constantes remodelações. Embora muitas retrocópias sejam elementos neutros ou disfuncionais, algumas desempenham papéis essenciais, seja na origem de novos genes ou na predisposição a doenças. O impacto dessas duplicações na diversidade biológica e na adaptação evolutiva reforça a importância de investigar seus mecanismos de origem, manutenção, evolução e suas consequências funcionais para as espécies, incluindo nós humanos.
Compreender os doppelgängers genômicos não é apenas um exercício conceitual, mas uma necessidade para avançarmos na biologia evolutiva e médica. O estudo das retrocópias nos permite enxergar como o genoma se reinventa, criando novas possibilidades funcionais e, ao mesmo tempo, carregando riscos inerentes a essa plasticidade. Assim, essas cópias continuam a moldar a trajetória da vida, ora como sombras genéticas, ora como protagonistas da inovação biológica.
Referências
- ALBERTS, B. et al. DNA e cromossomos; Do DNA ao RNA; Do RNA à proteína. In: Biologia molecular da célula. 6. ed. Porto Alegre: Artmed, 2017. p. 323–451.
- BURKI, F.; KAESSMANN, H. Birth and adaptive evolution of a hominoid gene that supports high neurotransmitter flux. Nat. Genet., v. 36, n. 10, p. 1061–1063, Oct. 2004.
- COST, G. J. et al. Human L1 element target-primed reverse transcription in vitro. EMBO J., v. 21, n. 21, p. 5899–5910, Nov. 2002.
- DAWKINS, R. O gene egoísta. Tradução: Rejane Rubino. São Paulo: Companhia das Letras, 2007.
- GAIMAN, N. Lugar nenhum. Tradução: Fábio M. Barreto. Rio de Janeiro: Intrínseca, 2016.
- GARZA, R. et al. LINE-1 retrotransposons drive human neuronal transcriptome complexity and functional diversification. Sci. Adv., v. 9, n. 44, p. eadh9543, Nov. 2023.
- HAN, J. S.; BOEKE, J. D. LINE-1 retrotransposons: modulators of quantity and quality of mammalian gene expression? Bioessays, v. 27, n. 8, p. 775–784, Aug. 2005.
- KAESSMANN, H.; VINCKENBOSCH, N.; LONG, M. RNA-based gene duplication: mechanistic and evolutionary insights. Nat. Rev. Genet., v. 10, n. 1, p. 19–31, Jan. 2009.
- MARTIN, S. L. et al. The structures of mouse and human L1 elements reflect their insertion mechanism. Cytogenet. Genome Res., v. 110, n. 1-4, p. 223–228, 2005.
- MARTIN, S. L. Nucleic acid chaperone properties of ORF1p from the non-LTR retrotransposon, LINE-1. RNA Biol., v. 7, n. 6, p. 706–711, Nov. 2010.
- OGINO, S. et al. A cohort study of tumoral LINE-1 hypomethylation and prognosis in colon cancer. J. Natl. Cancer Inst., v. 100, n. 23, p. 1734–1738, Dec. 2008.
- POLISENO, L. et al. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature, v. 465, n. 7301, p. 1033–1038, Jun. 2010.
- SULTANA, T. et al. Integration site selection by retroviruses and transposable elements in eukaryotes. Nat. Rev. Genet., v. 18, n. 5, p. 292–308, May 2017.
- SUN, Z. et al. LINE-1 promotes tumorigenicity and exacerbates tumor progression via stimulating metabolism reprogramming in non-small cell lung cancer. Mol. Cancer, v. 21, n. 1, p. 147, Jul. 2022.
- SZAK, S. T. et al. Genome Biol, v. 3, n. 10, p. research0052.1, 2002.
- TELESNITSKY, A.; GOFF, S. P. Reverse transcriptase and the generation of retroviral DNA. In: Retroviruses. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1997.
- VANIN, E. F. Processed pseudogenes: characteristics and evolution. Annu. Rev. Genet., v. 19, n. 1, p. 253–272, 1985.
- VASCONCELOS, F. T. G. R. D. E. et al. Revisitando o Dogma Central. Genet. Esc., v. 16, n. 2, p. 196–207, Jun. 2021.
- VIOLLET, S.; MONOT, C.; CRISTOFARI, G. L1 retrotransposition: The snap-velcro model and its consequences. Mob. Genet. Elements, v. 4, n. 1, p. e28907, Jan. 2014.
- WEI, W. et al. Human L1 retrotransposition: cis preference versus trans complementation. Mol. Cell. Biol., v. 21, n. 4, p. 1429–1439, Feb. 2001.
- VAN HOESEL, A. Q. et al. Hypomethylation of LINE-1 in primary tumor has poor prognosis in young breast cancer patients: a retrospective cohort study. Breast Cancer Res. Treat., v. 134, n. 3, p. 1103–1114, Aug. 2012.
- VON GOETHE, J. W. Os sofrimentos do jovem Werther. Tradução: Claudia Cavalcanti. São Paulo: Martin Claret, 2014.
MILLER, Thiago L. A. Espelhos do genoma: retrocópias como doppelgängers moleculares. Galantelab, 5 de fevereiro de 2025. Disponível em: https://galantelab.github.io/blog/dna/l1/elementos%20transpon%C3%ADveis/retroc%C3%B3pia/2025/02/05/retrocopias-como-doppelgangers-moleculares.html. Acesso em:
 « Genes saltadores e a resposta à cisplatina em câncer de ovário
« Genes saltadores e a resposta à cisplatina em câncer de ovário
 Desvendando o mundo surpreendente das retrocópias »
Desvendando o mundo surpreendente das retrocópias »

Thiago L. A. Miller